
- jaderná blána je tvořena dvěma listy
- spoje jaderné blány tvoří póry
- vnitřní obsah sestává z DNA, RNA a proteinů
Komplex těchto molekul tvoří rozptýlenou hmotu, tzv. chromatin. Součástí jádra je také snadno pozorovatelné jadérko (nucleolus).
Všechno jen ne se šprtat!
Komplex těchto molekul tvoří rozptýlenou hmotu, tzv. chromatin. Součástí jádra je také snadno pozorovatelné jadérko (nucleolus).
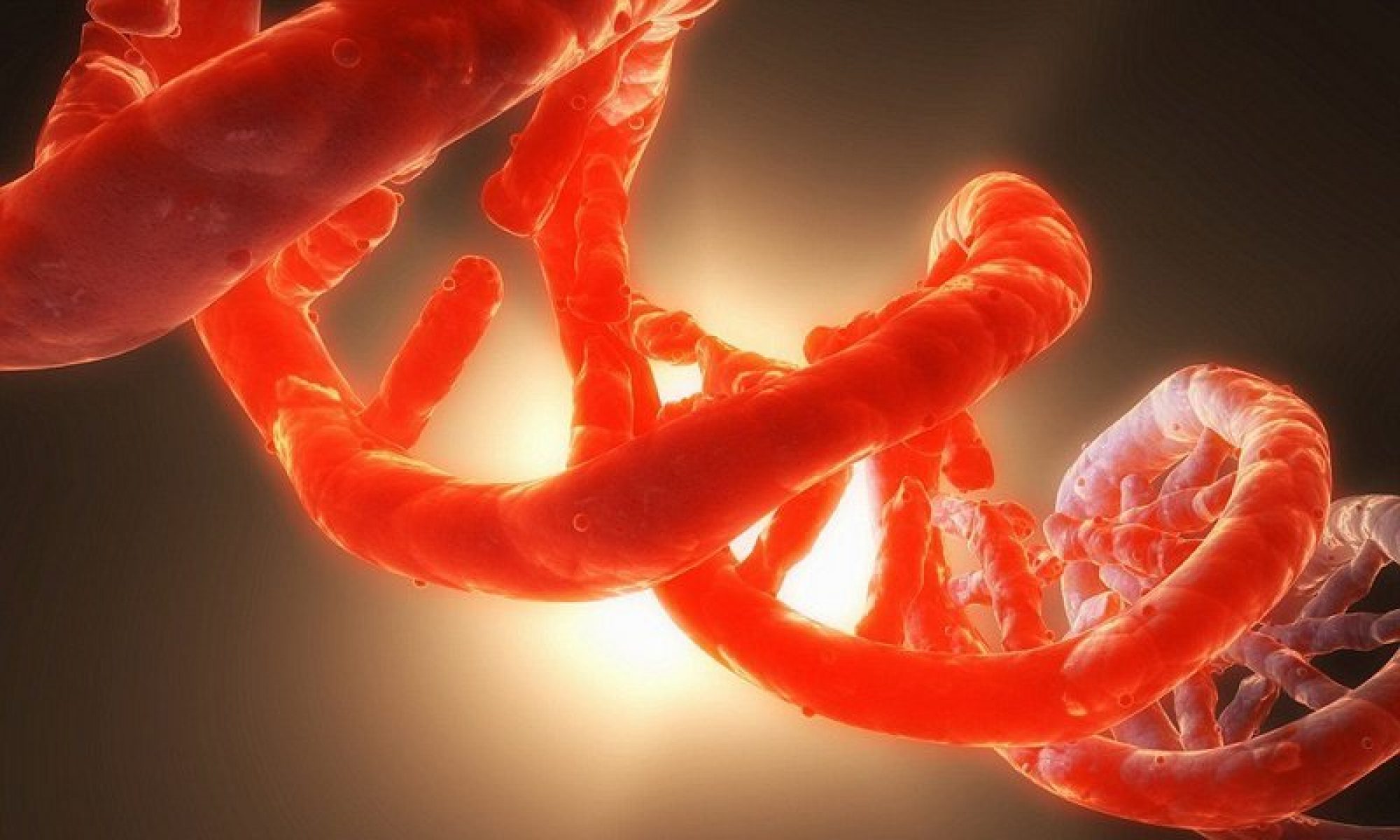
Při vzniku pohlavních buněk se rozchází do pohlavních buněk vždy po jedné alele z párů – tak u monohybrida vznikající 2 typy gamet, jsou vůči sobě v poměru 50:50 při zachování pravidla o volné kombinovatelnosti genů-jednoho z Mendelových zákonů.
Buněčný cyklus má 5 fází:
G (0,1), S, G2 a M.
M fáze. Je poslední etapou buněčného cyklu.
Mitóza. Je vlastní dělení buňky. Probíhá ve 4 fázích: Profáze, metafáze, anafáze a telofáze.
Meióza. Meióza neboli zrací dělení má funkci při redukci diploidu na haploid. Meióza se dělí na dvě základní etapy:
Meióza I a Meióza II. Ty se dále dělí:
Meióza I – má 3 fáze:
Profáze I a ta se dále dělí na:
Metafáze I
Anafáze I
Meióza II: Toto dělení je podobné mitóze (profáze II).
a) Struktura NK
Základními stavebními kameny obou kyselin (DNA/RNA) jsou mononukleotidy, složené z pentózy (C=5), z fosfátu a dusíkaté báze. Spojení je zajištěno fosfodiesterovou vazbou a to kovalentně. Sacharidem může být buď deoxyribóza (2´-deoxy-D-ribofuranóza), nebo u RNA ribóza (D-ribofuranóza). Teprve napojením kyseliny trihydrogenfosforečné na nukleozid (popisované v předchozím kroku) vzniká nukleotid. Existuje pět základních bází – A, G, T, C, U A=Adenin G=Guanin T=Thymin C=Cytosin U=Uracil RNA je tvořena 4-mi z těchto pěti základních bází (A, U, C, G) DNA je tvořena taktéž 4-mi z těchto pěti základních bází (A, T, C, G)b) Tvar NK
Fosfodiesterovými vazbami spojené mononukleotidy tvoří oligonukleotidové nebo dlouhé polynukleotidové řetězce, kde fosfát vždy váže 5´konec jedné pentózy s 3´koncem následujícího cukru. Syntéza, kterou zprostředkují enzymy DNA nebo RNA polymerázy (podle toho o jakou NK se jedná), tedy probíhá směrem od konce 5´ke konci 3´. Variabilita v primární struktuře DNA a RNA je založena na zmíněném střídání bází v řetězci. Druhotným tvarem polynukleotidových DNA řetězců je šroubovice, nejčastěji dvoušroubovice. NK jsou hlavní složkou jádra každé buňky. Základní tvar DNA je pravotočivá dvoušroubovice. Adenin se páruje dvěma vodíkovými můstky s thyminem a guanin s cytozinem třemi. Purinová a pyrimidinová báze se tedy páruje. Párování jednotlivých bází se nazývá komplementarita.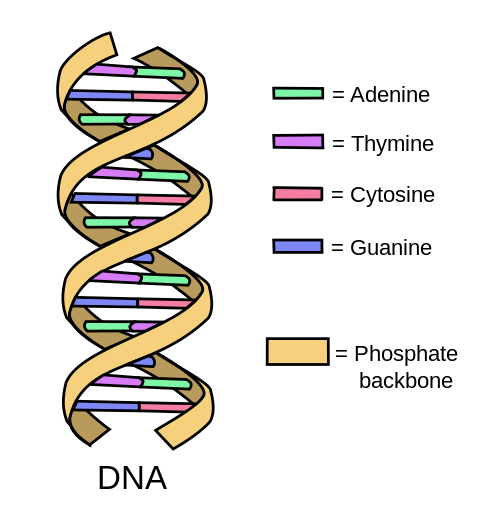
Základní schéma DNA > RNA > protein a zvláštní případ RNA > DNA > RNA > protein. Podle současných představ můžeme strukturní gen definovat jako sekvenci nukleotidů DNA (příp. RNA) s informací pro 1 polypeptidový řetězec (bílkovinu).
DNA = deoxyribonukleová kyselina, RNA = ribonukleová kyselina (1-vlákenná)
Podle předlohy DNA nebo syntéza RNA probíhá podle RNA předlohy, jedná se o replikaci (kopírování, ale nově vzniklý řetězec není zcela totožný). Nově vzniklý řetězec je komplementární, ale má obrácenou polaritu. Proces předchází dělení jakékoli buňky. Ori je místo začátku replikace, která probíhá oběma směry.
2.7.2 Průběh replikace
2.7.3 Transkripce
Syntézu RNA, podle předlohy DNA katalyzují na DNA závislé RNA polymerázy. Proces prodlužování řetězce a směr syntézy je stejný jako při replikace DNA. Zde je nutné nastudovat strukturu vlákna mRNA viz. obrázek.

Na rozdíl od replikace je přepis do RNA nesymetrický, v určitém segmentu DNA je přepisováno jen jedno vlákno směrem od 3´konce k 5´konci označované jako negativní (antikódující). Druhé nepřepisované vlákno (pozitivní) má potom stejnou sekvenci jako produkt transkripce, který je označován jako primární transkript. Schéma procesu je tedy následující:
pozitivní vlákno > přepisovaný řetězec > (transkripce) > RNA primární transkript
Průběh transkripce
a) Iniciace
b) Prodlužování transkriptu
c) Ukončení transkripce
d) Úpravy mRNA
2.7.4 Překlad genetické informace a proteosyntáza
Proce překladu „jazyka“ nukleotidů do pořadí aminokyselin (či jinak AMK) v polypeptidovém (bílkovinném) řetězci probíhá v buněčných organelách – ribozómech. Oproti replikaci/ transkripci je translace daleko složitější.
Průběh translace
a) Aktivace aminokyselin
b) Iniciace translace
c) Elongace peptidového řetězce
d) Ukončení translace
2.8 Genetický kód a jeho vlastnosti
Genetický kód je soubor pravidel, podle nichž je informace uložená v pořadí nukleotidů v mRNA (DNA) přeložena do primární struktury polypeptidového (bílkovinného) řetězce.
Kód má následující vlastnosti:
a) Je 3-písmenový/ tripletový
Základní jednotkou je trojice nukleotidů (triplet, kodón), která při translaci kóduje jednu AMK. Kodóny jsou čteny na ribozómu ve směru 5´> 3´. Existují 3 “ Stop kodóny“ a dále jeden kodón startovní (AUG). Pokud je AUG na začátku řetězce řídí methionin a zároveň zahajuje translaci, pokud je uprostřed, kóduje pouze AMK methionin.
b) Je nepřekrývající se
c) Je univerzální
d) Ja nadbytečný (redundantní, degenerovaný)
Většina z 20 AMK je kódována více než jedním tripletem nukleotidů (kodónem). Z výše uvedených vlastností existují výjimky. U některých bakterií a zejména u podbuněčných forem se čtení kódu může překrývat, tzn. posunem čtecího rámce jsou některé sekvence virů čteny i 3x (!) za vzniku různých funkčních polypeptidických řetězců. Viry takto efektivněji využívají svoji malou genetickou kapacitu (pro vznik řádově jednotek až desítek bílkovin). Např. v lidské mtDNA je terminační (ukončovací) kodón UGA čten jako tryptofan a izoleucinový kodón AUA je čten jako methionin. V rostlinné mtDNA triplet CGG nekóduje arginin, ale tryptofan. Další rozdíly existují u kvasinek a prvoků.
2.9 Struktura genomu (odkaz)
Genom prokaryontů
V porovnání s eukaryonty je maximálně využit. Více viz. skripta
Genom eukaryontů
Většina genů je ve více kopiích a tvoří genové rodiny. Jde o úzkou vazbu genů, které mají příbuznou stavbu a produkty které řídí, mají podobné funkce. Vznikly genovými duplikacemi (multiplikacemi) a nerovnoměrným překřížením chromozómů během meiózy.
AGA28E Biometrická genetika – Mgr. – ZS 2020/2021 – FAPPZ
AGA25E Cytogenetika a karyologie – Mgr. – ZS 2020/2021 – FAPPZ
AGA88E Genetika a šlechtění zvířat – Mgr. – ZS 2020/2021 – FAPPZ
AGA31E Genetika a šlechtění zvířat – Mgr. – ZS 2020/2021 – FAPPZ
2020/2021 – doktorská studia – FLD
2020/2021 – doktorská studia – FLD
AGA21E Genetika rostlin – Mgr. – LS 2020/2021 – FAPPZ
DAGX03Y Genetika zvířat – ZS 2020/2021 – doktorská studia – FAPPZ
DAGX03Y Genetika zvířat – LS 2020/2021 – doktorská studia – FAPPZ
DAGA04Y Molekulární genetika – ZS 2020/2021 – doktorská studia – FAPPZ
DAGA04Y Molekulární genetika – LS 2020/2021 – doktorská studia – FAPPZ
DAGX02Y Molekulární genetika a šlechtění rostlin – ZS 2020/2021 – doktorská studia – FAPPZ
DAGX02Y Molekulární genetika a šlechtění rostlin – LS 2020/2021 – doktorská studia – FAPPZ
DAGA07Y Molekularní genetika speciální – ZS 2020/2021 – doktorská studia – FAPPZ
DAGA07Y Molekularní genetika speciální – LS 2020/2021 – doktorská studia – FAPPZ
AGA70E Molekulární genetika zvířat – ZS 2020/2021 – FAPPZ
AGA35E Molekulární genetika zvířat – ZS 2020/2021 – FAPPZ
AGA01E Obecná genetika – ZS 2020/2021 – FAPPZ
AGA72E Obecná genetika – ZS 2020/2021 – FAPPZ
AGI02E Obecná genetika – Bc. – ZS 2020/2021 – FAPPZ
AGA74E Obecná genetika – HU – Bc. – LS 2020/2021 – FAPPZ
ALI01E Obecná zootechnika a genetika – Bc. – ZS 2020/2021 – FAPPZ
Postihuje četnost výskytu náhodného jevu v n nezávislých pokusech. Příklad: pravděpodobnost, že se narodí chlapec či dívka u rodičů se 2, 3, 4,…,n dětmi. Pokud speciálně n=1, jde o alternativní rozdělení. Binomické rozdělení se vyznačuje právě n pokusy.
Pokud se o inteligenci bavíme jako o kvantitativním znaku, pak její rozdělení v populaci odpovídá normálnímu (Gaussovu) rozdělení popsanému Gaussovou křivkou.

V genetice pracujícími s dvěma základními zdroji variability: genetické a vlivu prostředí, se upřednostňuje použití binomického rozdělení nad např. normálním.
Pro aplikaci binomického rozdělení je třeba znát parametry binomického rozdělení „n“ a „p“, hodnota q je dána vztahem q = 1 – p neboli závisí na p.
Důležitým poznatkem je, že výsledkem genetického procesu nemusí být jen dva alternativní výsledky. Genotypy a fenotypy ve štěpícím potomstvu nabývají více hodnot.
Později se lze setkat se znázorněním binomického rozdělení v potomstvech jako odezvy na selekci – genetický zisk (kapitola 13.2).